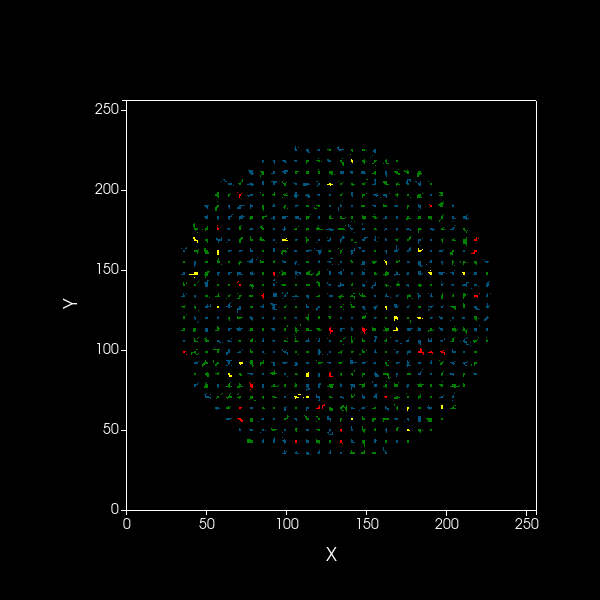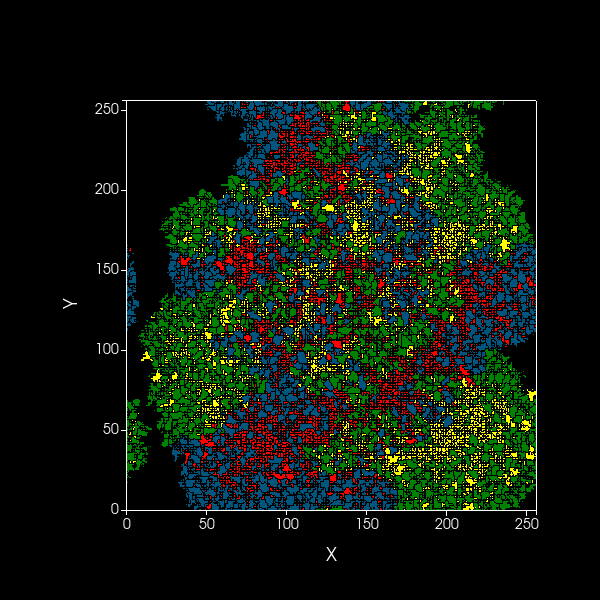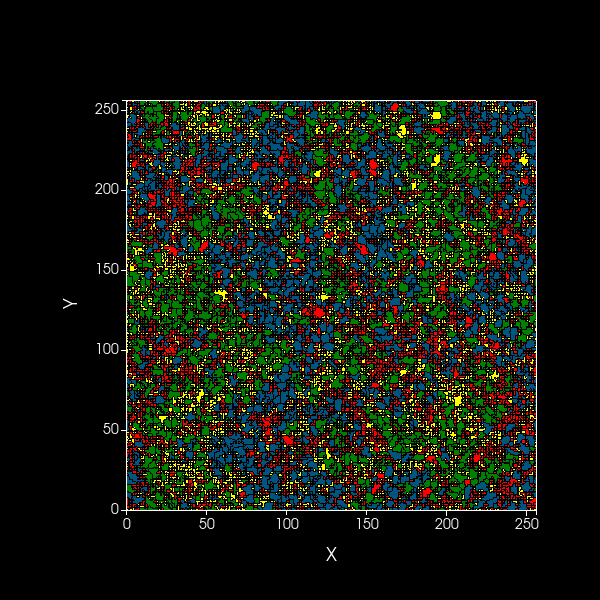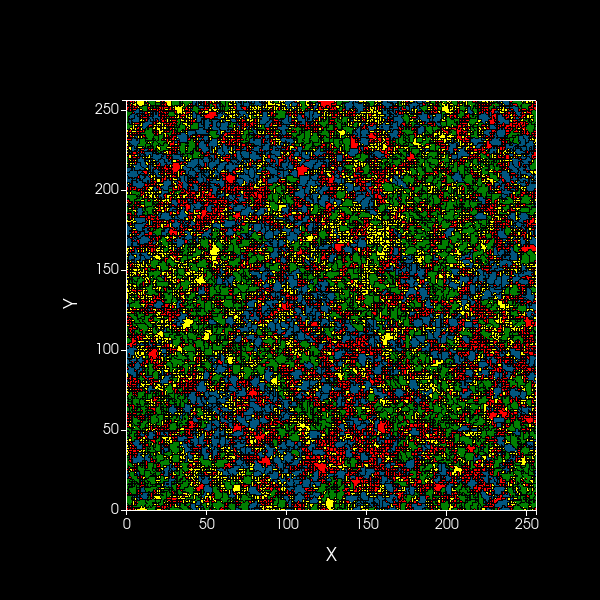
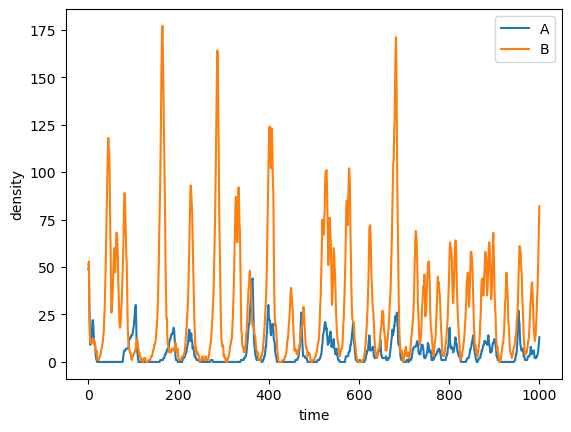
Simulation of spatial model showing dynamics over 1000 timesteps of A (blue) and B (green) cooperators,
as well as dormant A (red) and B (yellow) cells, on a 2D plane.
Black edges are present to distinguish between cells.
Interactions between species or strains can be considered mutualistic when each species
benefits from the presence of the other. Mutualisms are very common in nature, such as
the relationship between plants and their insect pollinators. In bacteria, one form of
mutualism occurs when different strains produce resources that can be used by different strains.
However, while receiving benefits from a mutualistic partner can increase growth of an
organism, producing the benefits for that partner might be costly. These costs may inhibit
the evolution of these mutualisms or perhaps lead to cheating, where some individuals receive
the benefits of the mutualism without producing any costly resources themselves.
Dormancy, where certain individuals become inactive for some time, may help to stabilize
these relationships. For instance, while the costs of mutualism may lead to a disadvantage
when resources are abundant, the mutualism may still be beneficial when resources are scarce.
A seed bank of dormant cells may allow mutualistic cells to persist when selection favors cheaters until
periods where it favors cooperation. Additionally, environmental stochasticity can lead to
extinction of one of the two partners by chance. A seed bank can drastically reduce this
probability by maintaining a reserve of individuals of both partners.
To model situations where dormancy can help maintain cooperation, I developed two
individual-based models (IBMs): one non-spatial, and one spatial. Both implement cells as
the primary individuals, or agents, and these cells can be of two types existing in a single
population. They both share a common resource and produce secondary resources that can only
be used by the other cell type. Both implement stochastic processes and allow individuals to
make decisions based upon their current attributes. For more information about implementation,
please go to the project GitHub repo.
These models have shown that when two mutualistic species are capable of dormancy, they can
coexist in the long-term despite extreme resource limitation. Normally, without dormancy, neither
species can survive, and without cooperation, one strain will go extinct. With both dormancy
and mutualism, both are maintained. In the future, both models will require thorough parameter
sweeps to identify precise parameters where dormancy increases the probability of coexistence.
Despite long periods of energy limitation, dormancy helps mutualist survive and coexist in the long-term.
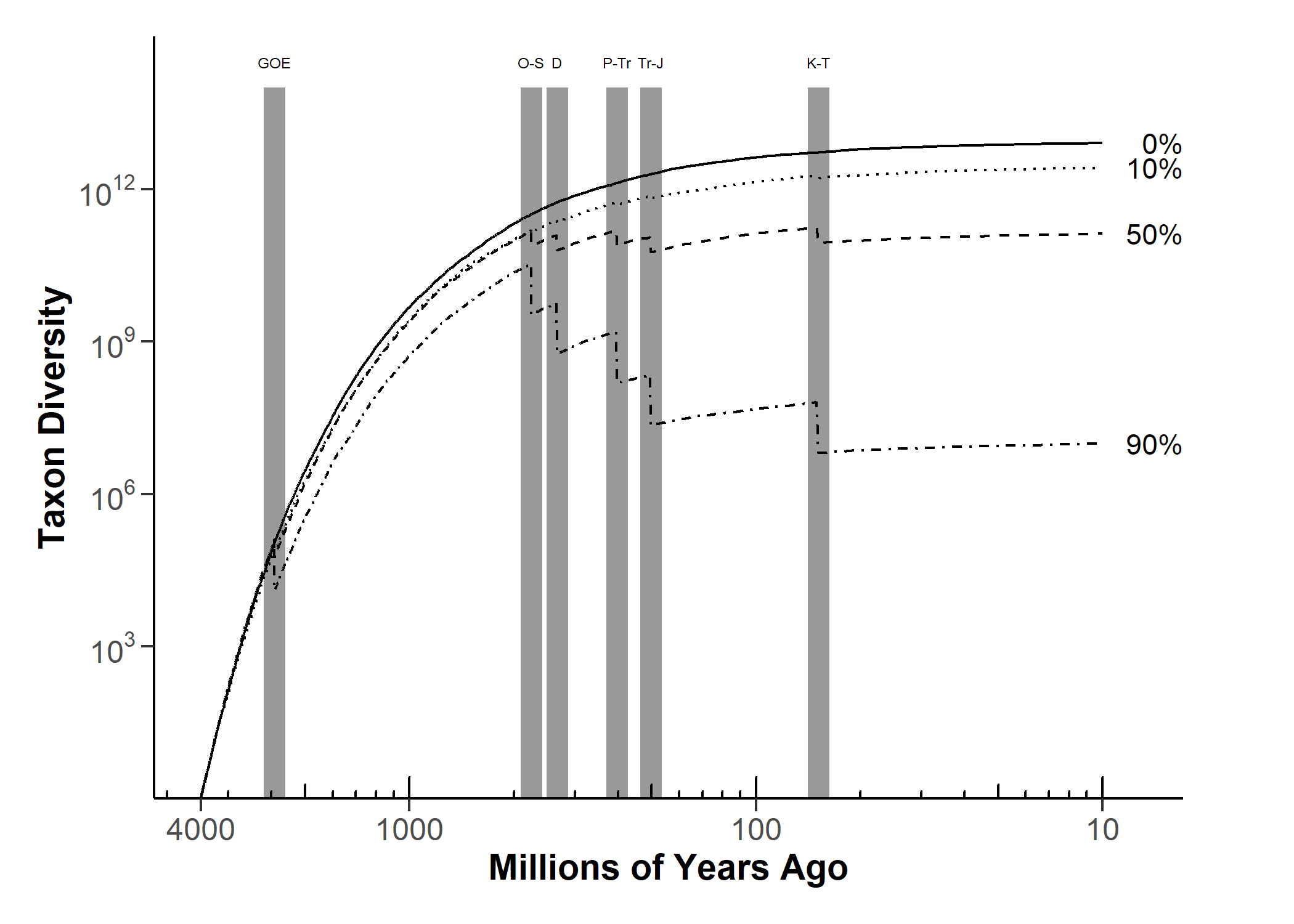
Simulations showing possible effects of mass extinction events (solid vertical bars) on microbial diversity.
Mass extinction here removes 0% (solid), 10% (dotted), 50% (dashed), or 90% (dot-dashed) of all species in that timestep.
GOE: Great Oxidation Event ; O-S: Ordovician-Silurian (445 Mya); D: Devonian (375 Mya); P-Tr: Permian-Triassic (252 Mya);
Tr-J: Triassic-Jurassic (201 Mya); K-T: Cretaceous (66 Mya).
It is difficult enough to count up all of the animal and plant species on the planet.
However, this task is even more challenging for microbes. Most bacteria that can be
detected from DNA sequencing approaches cannot even be grown in the lab, and the vast
majority of the globe has not been properly sequenced for its microbial content. While
several attempts have been made to estimate the extent of present-day microbial diversity,
the estimations range from 106 to 1012, and there is no way to
directly test these estimates.
One indirect way is to check the feasibility of each estimation. In order for there to
be a trillion microbial species, there must have been enough time passed to generate this
number of species. This is controlled by the speciation rate, the number of new species
formed per million years, and the extinction rate, the number of species that go extinct
per million years. The net result of speciation and extinction is known as the diversification
rate. To reach a trillion species after 4 billion years, the diversification rate must be high enough.
The converse is also true: if the diversification rate is too high, a million species might
be too low of an estimate.
With this logic, I used estimates of diversification rates of bacteria to fit stochastic birth-death models.
As there is no direct fossil record of bacteria, these estimates were taken from the
symbiotic bacteria of insects that had fossil records themselves. I also used models that included
global mass extinction events. Our results found that while the outcome of millions of species
is marginally more likely than billions or trillions, many combinations of parameters can lead
to even higher than trillions of species, even when accounting for mass extinction events.
This study is currently being written up into a manuscript for submission for publication.
Bacteriophages infecting E. coli. Credits Eye of Science.
Like all organisms, bacteria deal with infections from viruses. These antagonistic
interactions with bacteria-infecting viruses, also called bacteriophages or phages,
decrease the survival of bacteria. To limit these negative interactions, bacteria must
evolve ways to defend themselves and resist phage infection. This resistance can manifest
itself in several forms: modifying the surfaces to which phages need to attach, destroying
invasive DNA or RNA once inside the host or even early cell death to prevent larger outbreaks.
These mechanisms are only a small portion of the vast array of ways bacteria can defend
themselves against phages. Why do so many mechanisms exist? One potential explanation is
the other side of this picture: phage evolution. Phage mutations that can overcome the
defense mechanisms of their bacterial hosts confer advantages to these new mutants.
These new mutations can lead to new responses in the hosts to combat these new and improved
phages, thus starting a cycle of coevolution between host and virus.
This coevolution is the focus of my eco-evolutionary simulation model currently called
CommunityImmunity.
In this model, host and phage strains are allowed to mutate and create new strains that
may be phage-resistant or new phages that may have a larger host range. The demographic
information of all strains is tracked and partially determines the frequency of mutation.
I am currently using this model to explore the impact of increasing host and phage mutation
rate on the outcome of each simulation and the mode of coevolution.